Viruses are characterized by the following features:
(1) Viruses are particles composed of an internal core containing either DNA or RNA (but not both)
(2) Viruses must reproduce (replicate) within cells, viruses are obligate intracellular parasites. (The only bacteria that are obligate intracellular parasites are chlamydiae and rickettsiae.)
(3) Viruses replicate in a manner different from that of cells
SIZE & SHAPE OF VIRUSES
The shape of virus particles is determined by the arrangement of the repeating subunits/capsomere that form the protein coat (capsid)
Each capsomer consists one or several proteins
VIRAL NUCLEIC ACIDS
Only viruses have genetic material composed of single stranded DNA or of single-stranded or double-stranded RNA. The nucleic acid can be either linear or circular. The DNA is always a single molecule; the RNA can exist either as a single molecule or in several pieces. For example, both influenza virus and rotavirus have a segmented RNA genome. Almost all viruses contain only a single copy of their genome (i.e., they are haploid). The exception is the retrovirus family, whose members have two copies of their RNA genome (i.e., they are diploid).
Nucleocapsid – the capsid of a virus with the enclosed nucleic acid
Viral nucleocapsids have two forms of symmetry:
(1) Icosahedral, in which the capsomers are arranged in 20 triangles that form a symmetric figure (an icosahedron) with the approximate outline of a sphere;
(2) Helical, in which the capsomers are arranged in a hollow coil that appears rod-shaped. The helix can be either rigid or flexible. All human viruses that have a helical nucleocapsid are enclosed by an outer membrane called an envelope (i.e., there are no naked helical viruses). Viruses that have an icosahedral nucleocapsid can be either enveloped or naked
The advantage of building the virus particle from identical protein subunits is twofold:
(1) It reduces the need for genetic information, and
(2) It promotes self-assembly (i.e., no enzyme or energy is required).
VIRAL PROTEINS
The capsid proteins protect the genome DNA or RNA from degradation by nucleases. The proteins on the surface of the virus mediate the attachment of the virus to specific receptors on the host cell surface. This interaction of the viral proteins with the cell receptor is the major determinant of species and organ specificity. Outer viral proteins are also important antigens that induce neutralizing antibody and activate cytotoxic T cells to kill virus-infected cells.
The term “serotype” is used to describe a subcategory of a virus based on its surface antigens. For example, measles virus has one serotype, polioviruses have three serotypes, and rhinoviruses have over 100 serotypes. This is because its surface protein that induces neutralizing antibody capable of preventing infection. In contrast, polioviruses have three different antigenic determinants on its surface proteins, i.e., poliovirus type 1 has one kind of antigenic determinant, poliovirus type 2 has a different antigenic determinant, and poliovirus type 3 has a different antigenic determinant from types 1 and 2, hence polioviruses have three serotypes. There are two important medical implications of this, one, is that a person can be immune (have antibodies) to poliovirus type 1 and still get the disease, poliomyelitis caused by poliovirus types 2 or 3. The other implication is the polio vaccine must contain all three serotypes in order to be completely protective.
VIRAL ENVELOPE
Enveloped viruses are more sensitive to heat, drying, detergents, and lipid solvents such as alcohol and ether than are nonenveloped (nucleocapsid) viruses.
The surface proteins of the virus, whether they are the capsid proteins or the envelope glycoproteins, are the principal antigens against which the host mounts its immune response to viruses.
ATYPICAL VIRUS LIKE AGENTS
(1) Defective viruses are composed of viral nucleic acid and proteins but cannot replicate without a “helper” virus, which provides the missing function. Defective viruses usually have a mutation or a deletion of part of their genetic material
(2) Pseudovirions contain host cell DNA instead of viral DNA within the capsid. They are formed during infection with certain viruses when the host cell DNA is fragmented and pieces of it are incorporated within the capsid protein. Pseudovirions can infect cells, but they do not replicate.
(3) Viroids consist solely of a single molecule of circular RNA without a protein coat or envelope. Not in human disease
(4) Prions are infectious particles composed entirely of protein. They have no DNA or RNA.
When these proteins are in the normal, alpha-helix configuration, they are nonpathogenic, but when their configuration changes to a beta-pleated sheet, they aggregate into filaments, which disrupts neuronal function and results in the symptoms of disease.
Prions are highly resistant to inactivation by ultraviolet light, heat, and other inactivating agents. As a result, they have been inadvertently transmitted by human growth hormone and neurosurgical instruments.
Because they are normal human proteins, they do not elicit an inflammatory response or an antibody response in humans.
SPECIFIC EVENTS DURING THE GROWTH CYCLE
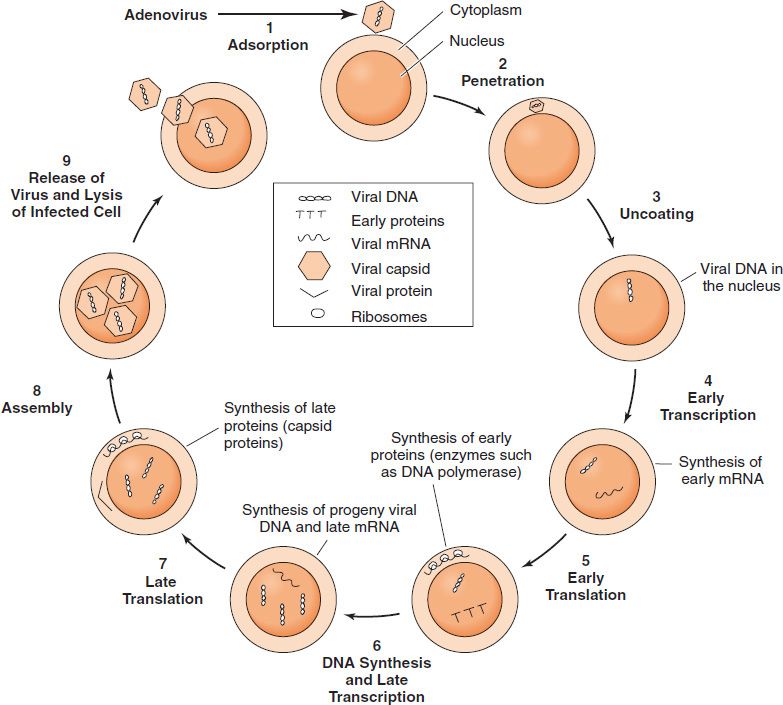
- The infecting parental virus particle attaches to the cell membrane and then penetrates the host cell.
- The viral genome is“uncoated” by removing the capsid proteins, and the genome is free to function.
- Early mRNA and proteins are synthesized; the early proteins are enzymes used to replicate the viral genome.
- Late mRNA and proteins are then synthesized. These late proteins are the structural, capsid proteins.
- The progeny virions are assembled from the replicated genetic material, and newly made capsid proteins and are then released from the cell
Attachment, Penetration, & Uncoating
The proteins on the surface of the virion attach to specific
receptor proteins on the cell surface through weak, noncovalent
bonding.
The specificity of attachment determines the host range of the virus
For example, herpes simplex virus type 1 attaches to the fibroblast growth factor receptor, rabies virus to the acetylcholine receptor, and human immunodeficiency virus (HIV) to the CD4 protein on helper T lymphocytes
The virus particle penetrates by being engulfed in a pinocytotic vesicle, within which the process of uncoating begins. A low pH within the vesicle favors uncoating.
Rupture of the vesicle or fusion of the outer layer of virus with the vesicle membrane deposits the inner core of the virus into the cytoplasm.
Gene Expression & Genome Replication
The first step in viral gene expression is mRNA synthesis.
DNA viruses, with one exception, replicate in the nucleus and use the host cell DNA-dependent RNA polymerase to synthesize their mRNA.
The poxviruses are the exception because they replicate in the cytoplasm, where they do not have access to the host cell RNA polymerase. They therefore carry their own polymerase within the virus particle.
The genome of all DNA viruses consists of double- stranded DNA, except for the parvoviruses, which have a single-stranded DNA genome
Most RNA viruses undergo their entire replicative cycle in the cytoplasm. The two principal exceptions are retroviruses and influenza viruses. Retroviruses integrate a DNA copy of their genome into the host cell DNA, and influenza viruses synthesize their progeny genomes in the nucleus. In addition, the mRNA of hepatitis delta virus is also synthesized in the nucleus of hepatocytes.
The genome of all RNA viruses consists of single stranded RNA, except for members of the reovirus family, which have a double-stranded RNA genome. Rotavirus is the important human pathogen in the reovirus family.
RNA viruses fall into four groups with quite different strategies for synthesizing mRNA:
(1) The simplest strategy is illustrated by poliovirus, which has single-stranded RNA of positive polarity1 as its genetic material. These viruses use their RNA genome directly as mRNA.
(2) The second group has single-stranded RNA of negative polarity as its genetic material. An mRNA must be transcribed by using the negative strand as a template. Because the cell does not have an RNA polymerase capable of using RNA as a template, the virus carries its own RNAdependent RNA polymerase. There are two subcategories of negative-polarity RNA viruses: those that have a single piece of RNA (e.g., measles virus [a paramyxovirus] or rabies virus [a rhabdovirus]) and those that have multiple pieces of RNA (e.g., influenza virus [a myxovirus]).
Certain viruses, such as arenaviruses and some bunyaviruses, have a segmented RNA genome, most of which is negative stranded, but there are some positive strand regions as well. RNA segments that contain both positive polarity and negative polarity regions are called “ambisense.”
(3) The third group has double-stranded RNA as its genetic material. Because the cell has no enzyme capable of transcribing this RNA into mRNA, the virus carries its own polymerase. Note that plus strand in double-stranded RNA cannot be used as mRNA because it is hydrogen-bonded to the negative strand. Rotavirus, an important cause of diarrhea in children, has 11 segments of double-stranded RNA.
(4) The fourth group, exemplified by retroviruses, has single-stranded RNA of positive polarity that is transcribed into double-stranded DNA by the RNA-dependent DNA polymerase (reverse transcriptase) carried by the virus.
This DNA copy is then transcribed into viral mRNA by the regular host cell RNA polymerase (polymerase II). Retroviruses are the only family of viruses that are diploid (i.e., that have two copies of their genome RNA).
These differences explain why some viruses yield infectious nucleic acid and others do not. Viruses that do not require a polymerase in the virion can produce infectious DNA or RNA. By contrast, viruses such as the poxviruses, the negative-stranded RNA viruses, the double-stranded RNA viruses, and the retroviruses, which require a virion polymerase, cannot yield infectious nucleic acid.
Once the viral mRNA of either DNA or RNA viruses is synthesized, it is translated by host cell ribosomes into viral proteins, some of which are early proteins (i.e., enzymes required for replication of the viral genome) and others of which are late proteins (i.e., structural proteins) of the progeny viruses. (The term early is defined as occurring before the replication of the genome, and late is defined as occurring after genome replication.) The most important of the early proteins for many RNA viruses is the polymerase that will synthesize many copies of viral genetic material for the progeny virus particles. No matter how a virus makes its mRNA, most viruses make a virus-encoded polymerase (a replicase) that replicates the genome (i.e., that makes many copies of the parental genome that will become the genome of the progeny virions).
Some viral mRNAs are translated into precursor polypeptides that must be cleaved by proteases to produce the functional structural proteins whereas other viral mRNAs are translated directly into structural proteins.
Replication of the viral genome is governed by the principle of complementarity, which requires that a strand with a complementary base sequence be synthesized; this strand then serves as the template for the synthesis of the actual viral genome.
(1) poliovirus makes a negative strand intermediate, which is the template for the positivestrand genome;
(2) influenza, measles, and rabies viruses make a positive-strand intermediate, which is the template for the negative-strand genome;
(3) rotavirus makes a positive strand that acts both as mRNA and as the template for the negative strand in the double-stranded genome RNA;
(4) retroviruses use the negative strand of the DNA intermediate to make positive-strand progeny RNA;
(5) hepatitis B virus uses its mRNA as a template to make progeny double stranded DNA;
(6) the other double-stranded DNA viruses replicate their DNA by the same semiconservative process by which cell DNA is synthesized
As the replication of the viral genome proceeds, the structural capsid proteins to be used in the progeny virus particles are synthesized
Assembly & Release
The progeny particles are assembled by packaging the viral nucleic acid within the capsid proteins. The action of enzymes and expenditure of energy are not required
Virus particles are released from the cell by either of two processes. One is rupture of the cell membrane and release of the mature particles; this usually occurs with unenveloped viruses. The other, which occurs with enveloped viruses, is release of viruses by budding through the outer cell membrane (An exception is the herpes virus family, whose members acquire their envelopes from the nuclear membrane rather than from the outer cell membrane.)
The budding process begins when virus-specific proteins enter the cell membrane at specific sites. The viral nucleocapsid then interacts with the specific membrane site mediated by the matrix protein. The cell membrane evaginates at that site, and an enveloped particle buds off from the membrane
LYSOGENY
- Lysogeny is the process by which viral DNA becomes integrated into host cell DNA, replication stops, and no progeny virus is made. Later, if DNA is damaged by, for example, UV light, viral DNA is excised from the host cell DNA, and progeny viruses are made. The integrated viral DNA is called a prophage.
Bacterial cells carrying a prophage can acquire new traits, such as the ability to produce exotoxins such as diphtheria toxin.
Transduction is the process by which viruses carry genes from one cell to another. Lysogenic conversion is the term used to indicate that the cell has acquired a new trait as a result of the integrated prophage.
Some viruses can use an alternative pathway, called the lysogenic cycle, in which the viral DNA becomes integrated into the host cell chromosome and no progeny virus particles are produced at that time viral nucleic acid continues to function in the integrated state in a variety of ways
Infection by lambda phage in E. coli begins with injection of the linear, double-stranded DNA genome through the phage tail into the cell. The linear DNA becomes a circle as the single-stranded regions on the ends pair their complementary bases. A ligating enzyme makes a covalent bond in each strand to close the circle. Circularization is important because it is the circular form that integrates into host cell DNA.
The choice between the 2 pathways depends on the balance between two proteins, the repressor produced by the c-I gene and the antagonizer of the repressor produced by the cro gene. If the repressor predominates, transcription of other early genes is shut off and lysogeny ensues.
The next important step in the lysogenic cycle is the integration of the viral DNA into the cell DNA. This occurs by the matching of a specific attachment site on the lambda DNA to a homologous site on the E. coli DNA and the integration (breakage and rejoining) of the two DNAs mediated by a phage-encoded recombination enzyme. The integrated viral DNA is called a prophage. Most lysogenic phages integrate at one or a few specific sites, but some, such as the Mu (or mutator) phage, can integrate their DNA at many sites, and other phages, such as the P1 phage, never actually integrate but remain in a “temperate” state extrachromosomally, similar to a plasmid
Because the integrated viral DNA is replicated along with the cell DNA, each daughter cell inherits a copy. However, the prophage is not permanently integrated. It can be induced to resume its replicative cycle by the action of UV light and certain chemicals that damage DNA. UV light induces the synthesis of a protease, which cleaves the repressor. Early genes then function, including the genes coding for the enzymes that excise the prophage from the cell DNA. The virus then completes its replicative cycle, leading to the production of progeny virus and lysis of the cell.
Relationship of Lysogeny in Bacteria to Latency in Human Cells
Shortly after HSV (herpes simplex virus) infects neurons, a set of “latency-associated transcripts” (LATS) are synthesized. These are noncoding, regulatory RNAs that suppress viral replication. The precise mechanism by which they do so is unclear. Reactivation of viral replication at a later time occurs when the genes encoding LATS are excised.
CMV employs different mechanisms. The CMV genome encodes microRNAs that inhibit the translation of mRNAs required for viral replication. Also, the CMV genome encodes both a protein and an RNA that inhibit apoptosis in infected cells. This allows the infected cell to survive.
NB:
- Infectious nucleic acid is viral genome DNA or RNA, purified free of all proteins, that can undergo the entire replicative cycle within a cell and produce infectious progeny viruses. Infectious nucleic acid, because it has no associated protein, can enter and replicate within cells that the intact virion cannot.
- Polarity of viral genome RNA: Genome RNA that has the same base sequence as the mRNA is, by definition, positivepolarity RNA. Most positive-polarity genomes are translated into viral proteins without the need for a polymerase in the virion. The exception is the retroviruses, which use reverse transcriptase in the virion to transcribe the genome RNA into DNA. Genome RNA that has a base sequence complementary to mRNA has, by definition, negative polarity. A virus with a negative-polarity RNA genome must have an RNA polymerase in the virion to synthesize its mRNA.
- Viral replication: All DNA viruses replicate in the nucleus, except poxviruses, which replicate in the cytoplasm. All RNA viruses replicate in the cytoplasm, except retroviruses, influenza virus, and hepatitis D virus, which require an intranuclear step in their replication. Many viruses encode a replicase, which is a DNA or RNA polymerase that synthesizes the many copies of the progeny viral genomes.
- Viral genome: The genome of all DNA viruses is doublestranded except for that of parvoviruses, which is singlestranded. The genome of all RNA viruses is single-stranded except for that of reoviruses (e.g., rotavirus), which is doublestranded.
